I had the opportunity to speak to Daniel Kolbin last week on WRFI 88.1FM's Locally Sourced Science Program (www.locallysourcedscience.org). This is a wonderful program that highlights some of the great science done here in Ithaca. We talked about Drosophila diversity and the Species Stock Center here at Cornell, as well as some of my research on Hawaiian Drosophila. Here's the audio file, with my great thanks to Daniel and the LSS program for the wonderful discussion!
|
Here's a short article on the Stock Center's new home at Cornell University. This news piece was posted in the Cornell Chronicle in late January. http://news.cornell.edu/stories/2018/01/lab-rats-molecular-biology-find-home-campus. The write up mainly talks about the living collection of ~300 species of Drosophila from all over the world. Our mission is to provide Drosophila cultures to researchers working on all aspects of Drosophila biology, from genetics and development to ecology and behavior. Thanks to reporter Krishna Ramanujan and photographer Chris Kitchen.
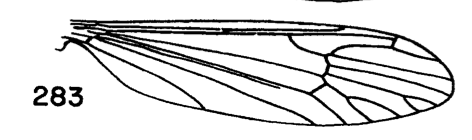
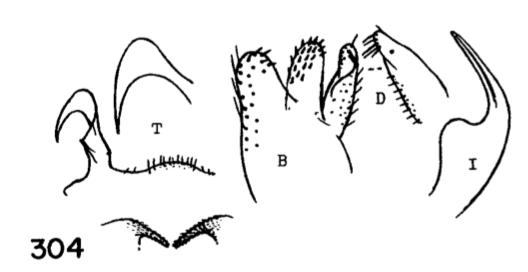
Dicranota Zetterstedt, 1838: 851The genus Dicranota is divided into 11 subgenera and includes about 280 species worldwide (Oosterbroek 2017). These include some of the smallest members of the family Pediciinae (Alexander 1967). The Nearctic fauna is comprised of 73 species placed in seven subgenera; Dicranota (15), Eudicranota (4), Paradicranota (4), Plectromyia (15), Polyangaeus (3), Rhaphidolabina (1), Rhaphidolabis (31). The subgenera were erected based on differences in wing venation, although there have been no phylogenetic studies aimed at testing the monophyly of either Dicranota or the included subgenera. Rhaphidolabis, the largest subgenus, seems to have a classic East/West split distribution, with seven species east of the Mississippi and the remaining 24 in the Western US. Alexander's (1967) treatment of the crane flies of California included a key to about 20 of the western species. Several species were excluded either because they were known only from females or were considered subspecies at the time. The updated key below includes all species west of the Mississippi, along with images from Alexander's descriptions (Alexander 1949, 1954, 1967). Key to the Western Species of Dicranota (Rhiphidolabis)1. General coloration of body yellow; male hypopygium with tergal arms produced caudad into long slender spines (Fig. 1)...xanthosoma Alexander, 1944. USA (Wash, Oreg). - General coloration of body brown or gray; male hypopygium not as above...2
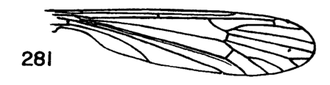
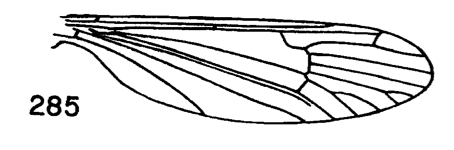
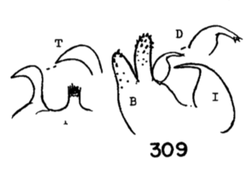
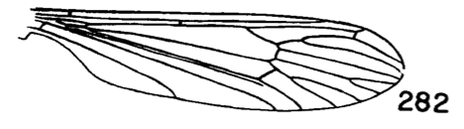 Figure 3. Wing of D. polymeroides, from Alexander (1967) 2. Antennae of male elongate, exceeding one-half the wing or extending to beyond base of abdomen; male hypopygium with posterior margin of tergite including broad obtuse lateral lobes and slender intermediate lobules that are separated by a broad U-shaped emargination (Fig. 2); venation (Fig. 3)...... ...polymeroides (Alexander, 1914). USA (Alaska, Wash, Idaho, south to Calif). - Antennae of male short, not extending to the wing root; male hypopygium with tergite not as above...3 3. Wings with Sc short, Sc1 ending opposite or before fork of Rs (e.g., Fig. 4)...4 - Wings with Sc longer, Sc1 ending some distance beyond fork of Rs, usually opposite or beyond the fork of R2+3+4...10 4. Wings with r-m ending before the fork of Rs, the latter very strongly arcuated, R2+3+4 short, suberect (Fig. 4)...tehama Alexander, 1950. USA (Calif). - Wings with r-m at or beyond fork of Rs, the latter relatively long, angulated and in cases short-spurred beyond origin (e.g., Fig. 5)...5
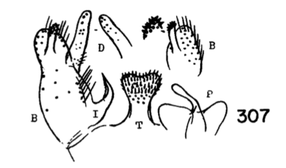
5. Wings with cell R3 petiolate; male hypopygium with tergal region not produced into lateral arms, the median region of posterior border with a small circular emargination behind which is a darkened area that bears numerous setae; basistyle without a modified arm on mesal face near apex (Fig. 6)............. ...fenderi Alexander, 1954. USA (Oreg). - Wings with cell R3 sessile; male hypopygium with posterior border of tergite broadly emarginate, the sides with pale strong arms (e.g., Fig. 7); basistyle on mesal face near apex with a short to longer pale arm that terminates in a blackened spine or elongate blackened blade, or else in a microscopic pale point (Fig. 8)...nooksackensis group, 6
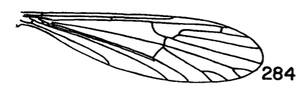
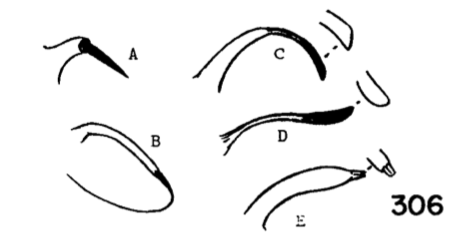
6. Apical arm of basistyle very short, terminating in a longer strong blackened acute spine (Fig. 8A)....... ...nooksackensis Alexander, 1949. Canada, USA (Alaska, south to Oreg and Calif). - Apical arm of basistyle longer, the pale base subequal to or longer than darkened outer part...7 7. Apical arm of basistyle stout, terminating in a microscopic pale point (Fig. 8E).................................... ...brevicula Alexander, 1966. USA (Calif). - Apical arm of basistyle terminating in a blackened blade or spot (Fig. 8B-D)...8 8. Apical arm of basistyle long and slender, terminating in a short blackened spinelike point that is extended farther into a paler setoid prolongation (Fig. 8B)...brevispinosa Alexander, 1958. USA (Wash). - Apical arm of basistyle shorter, terminating in a blackened blade, the apex obtuse...9 9. Apical arm of basistyle with the blackened apex longer, only slightly less than the pale basal stem (Fig 8C)...denningi Alexander, 1966. USA (Calif). - Apical arm of basistyle with the blackened apex relatively short, slightly more than one-half the pale basal stem (Fig. 8D)...evanescens Alexander, 1966. USA (Calif). 10. Wings with cell R3 sessile or subsessile, in the latter case (subsessilis) with R2+3+4 shorter than basal section of Rs, the latter equal to r-m (Figs. 9, 10)...11, sessilis (Alexander, 1917), USA (Colo), known from a single female, keys here. - Wings with cell R3 petiolate (Fig. 5)...16
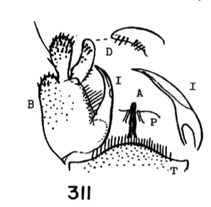
11. Male hypopygium (Fig. 11) with posterior border of ninth tergite broadly emarginate...12 - Male hypopygium (Figs. 12-14) with median region of posterior border of tergite slightly produced or truncate, not emarginate...13
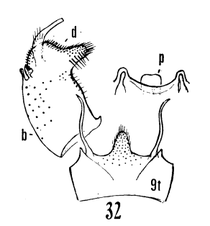
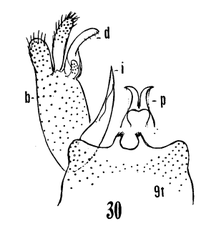
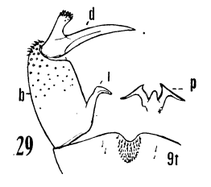
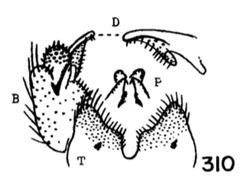
12. Size large (wing of 8 about 10 mm); wings with a brown cloud over r-m; basal section of vein Rs present, nearly as long as r-m; male hypopygium with the median emargination broad, the lobes divergent (Fig. 7)...uniplagia Alexander, 1954. USA (Oreg). While the wing was not drawn or described, the large body size and structure of the male hypopygium (Fig. 19), would place stigma (Alexander, 1924). USA (Wash, south to Colo), near this species. This may be incorrect if R3 is not sessile. - Size smaller (wing of 8 about 8 mm); wings without a darkened cloud over r-m; basal section of vein Rs obliterated, r-m at the fork of R4+5; male hypopygium (Fig. 11) with the median emargination the lobes less divergent...sanctaeluciae Alexander, 1964. USA (Calif). 13. Male hypopygium with median region of posterior border of tergite and mesal face of basistyle with unusually long setae (Fig. 12)...trichopyga Alexander, 1966. USA (Calif). - Male hypopygium (figs. 311,313) without unusually long setae on the tergite or basistyle (Figs, 14, 15)...14 14. Wings milky white, without stigmal darkening, veins pale (Fig. 10); male hypopygium with interbase a long swordlike blade; tergite without lateral arms (Figure 13)....................................................... ...subsessilis (Alexander, 1921). Canada, USA (BC to Mont and Wyo, south to Calif). - Wings with stigmal darkening more evident, veins darker; male hypopygium with interbase short, tip bispinous; lateral tergal arm present, their tips obtuse (Fig. 14)...15 15. Wings with stigma faint to scarcely &dent, in cases pale brown....................................................... ...cayuga (Alexander, 1916). Canada, USA (Alaska to Nfld, south to Oreg, Colo and SC). - Wings with stigma conspicuous, brown to dark brown (Fig. 9); male hypopygium (Fig. 14).............. ...vanduzeei (Alexander, 1930). USA (Calif). 16. Wings faintly suffused with brown, the stigma barely evident; R2+3+4 subequal to r-m (Fig. 5)..... ...nuptialis Alexander, 1948. USA (Calif). - Wings more whitened, stigma usually conspicuous; R2+3+4 slightly longer than r-m...17 17. Male hypopygium (Fig. 15) with basistyle produced into a single apical lobe, on its face with a slender lobule tipped with blackened spinoid setae; tergite with median region of posterior border produced into a conspicuous lobe that is weakly dilated a t outer end, the posterior border virtually truncate or very feebly emarginate...integriloba Alexander, 1943. USA (Wash and Idaho, south to Calif and Colo). - Male hypopygium (Figs. 16-18) with basistyle produced into two separated apical lobes; tergite with median region only slightly produced, the extreme in querula where the lobe is relatively short, the sides parallel...26
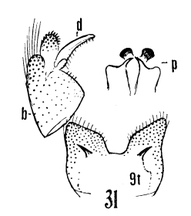
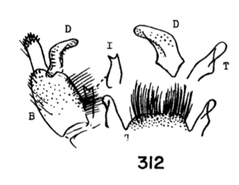
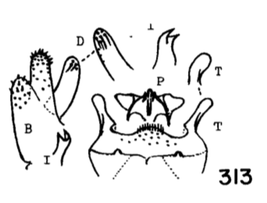
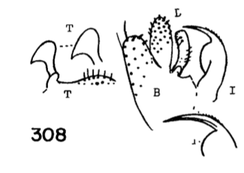
18. Wings with stigmal darkening very faint; male hypopygium with median region of tergite produced into a parallel-sided lobe; apex of dististyle narrowed into a slender lobe (Fig. 16)........................................ ...querula Alexander, 1944. USA (Oreg, Idaho, Utah and Calif). - Wings with stigmal darkening conspicuous; male hypopygium with median region of tergite convexly rounded; dististyle not narrowed at apex...19 19. Male hypopygium with lateral tergal lobes short and broad, the apical point short (Fig. 17).................. ...neomexicana (Alexander, 1912). Canada, USA (Alaska, south to Calif, Ariz, NMex and Colo). A former subspecies, subtruncata Alexander, 1949. USA (Wash), would key here. This species isn't illustrated but Alexander's description states "the ninth tergite virtually truncate, the margin at this point very slightly convex and with a concentration of setae. Two or three setae at the apex of the dististyle long and conspicuous." It should otherwise conform to D. neomexicana (Fig. 308). - Male hypopygium with lateral tergal lobes relatively narrow, extended into a long terminal spine (Fig. 18)...cazieriana Alexander, 1944. USA (Oreg, Calif and Utah).
Eastern Species (not included in key)
Dicranota (Rhaphidolabis) avis (Alexander, 1926). USA (Maine, south to NC). Dicranota (Rhaphidolabis) forceps (Alexander, 1924). Canada, USA (Mich to NSc, south to Tenn). Dicranota (Rhaphidolabis) hickmanae Alexander, 1940. USA (Tenn). Dicranota (Rhaphidolabis) persimilis (Alexander, 1920). USA (Pa and Md, south to Tenn and SC). Dicranota (Rhaphidolabis) rogersiana (Alexander, 1925). Canada, USA (Mich, Que). Dicranota (Rhaphidolabis) rubescens (Alexander, 1916). Canada, USA (Que to NSc, south to NY and Mass). Dicranota (Rhaphidolabis) tenuipes (Osten Sacken, 1869). Canada, USA (Mich to NBr, south to Md and Va). I'm going to summarize the collections that I made moving from Berkeley to Ithaca in summer 2017. This is going to be a work in progress but will *hopefully* soon contain lots of images, IDs and distributional information. It won't be posted chronologically but the order of states collected in was California -> Oregon -> Montana (no collections in Washington or Idaho) -> Wyoming.
And that was pretty much it for the western part of the trip. Once we got out of Wyoming, we were focused on driving to the east coast as quickly as possible. I'm sure there are lots of great craneflies in the intervening states (especially in the Black Hills of South Dakota) and I'm hoping to make trips in the future to discover them. The last state I have records for on this trip is Delaware, where we spent a week visiting Chelsea's parents before driving to Ithaca. Montana. I have only a single specimen from Montana, 1090.1. This was collected at a rest stop near Anaconda, MT. The specimen is a female and it has some minor damage. It is a species in the genus Limonia, although I'll need additional material before I can settle on a species ID. We drove through this state pretty quickly on our way to Yellowstone and I didn't have the time to survey or collect much. This will definitely require another trip! Wyoming. I wasn't able to spend very much time collecting in Wyoming. We were in Yellowstone for ~2 days and I didn't have permits. We did spend one night in the Big Horn Mountains on the eastern part of the state. This was an amazing campsite near Shell Creek. I managed to collect two different species at this locality (O'Grady 1091).
There were also two females, each representing a different species of the genus Tipula from this locality. Additional material will be needed before I can assign these to subgenus and species.
The National Drosophila Species Stock Center (DSSC) maintains about 2000 different stocks of ~250 individual Drosophila species. It was established in the 1940's by JT Patterson's laboratory at the University of Texas at Austin. It was maintained by Patterson's group, largely through the efforts of his former graduate student Marshall R. Wheeler, until 1982 when it moved to Bowling Green State University. Jong-Sik Yoon served as DSSC Director for the 18 years it was in Bowling Green. The DSSC moved to the University of Arizona in 2000 and Therese Markow became the Director. Dr. Markow moved the DSSC to the University of California, San Diego in 2008. This year the DSSC will move again, this time to Cornell University, where I will take over as Director. I am excited about having this amazing resource on campus and look forward to expanding the collection. The Department of Entomology is the home of the center and has provided an amazing space for the facility in Comstock Hall, named for entomologist John Henry Comstock. Renovations are ongoing but I'm going to share some recent photos here. I'll update them as equipment and flies arrive!
Comstock 162C is an office area and will be used for visiting researchers and material storage. Room 162D will be converted to a large refrigerator for storing fly food.
It's been a while since I've posted much on this site. Most of my attention has been focused on planning and executing the lab's move to the Department of Entomology at Cornell University. Graduate students Nina Pak and Allan Cabrero will be staying in Berkeley and I'll be admitting new students starting next fall.
 Crater Lake National Park We arrived in Portland where we were able to spend time with family (Bryonie, Eric, Oliver, and Guilyn) and friends (Susan and Jay). Paceyn had a wonderful time exploring the city. Her favorites were hanging with Oliver, shopping at Powell's Books, and taking the tram ride (twice) with Jay. Chelsea joined us on June 29th and we started our trip east.
Leaving Portland, we headed through Columbia River Gorge and ended up near Mt. Hood. Then it was through Washington and Montana to the north entrance to Yellowstone. Yellowstone was one of the highlights of the trip - what an amazing place!! We didn't get enough time to explore fully and are looking forward to another trip already.
We left the east entrance to Yellowstone and drove across Wyoming. The Big Horn Mountains and Shell Creek were incredible. We weren't expecting to find an area so beautiful and rich in fossils. All the roadcuts were even labeled by geological era. We left Wyoming and crossed over into the Black Hills and Mount Rushmore on the 4th of July. After that, we visited the Badlands, Wall Drug, and the Corn Palace.
We survived a tornado warning at a KOA in South Dakota (and Paceyn managed to make friends with a couple kids from San Francisco). After that, we spent another thousand or so miles of flat, boring states, many of which begin with an "I" (and which we didn't memorialize with photos). After a week visiting Chelsea's parents in Delaware, we drove to Ithaca and moved into our rental on the 15th of July.
The boxes are unpacked (mostly), the dust has settled (mostly), and "the lab" is now getting set up here in Ithaca, New York. There is still lots to do, including moving the National Drosophila Species Stock Center to its new home in the basement of Comstock Hall. I'll post more on that - and the flies I collected on the drive across country - later. Nina has been awarded a 2017 Rosemary Grant Graduate Student Research Award from the Society for the Study of Evolution and the Robert Usinger Award from ESPM. Both awards will go toward helping fund her research on the evolution of salt tolerance in Diptera. Well done!
https://nature.berkeley.edu/science-to-solutions/understanding-conserving-california-biodiversity-changing-climate/
We're still working out our lab meetings for this semester. Here's the tentative schedule:
2 October 9 October: Nina 16 October: Practice for Natalie’s talk at Southwest Fisheries Society 23 October 30 October: Grace and Patrick No lab meetings in November 4 December: Patrick |
PatrickProfessor Archives
April 2018
Categories
All
|






































 RSS Feed
RSS Feed
